Содержание
Механизм подкормок биопрепаратами
Биораствор питательных элементов – умная подкормка растений
Запас почвенных элементов питания в сильно ограниченном пространстве ящика очень быстро истощается. Рассада с каждым днём потребляет всё больше и больше макро- и микроэлементов, формируя свои ткани и органы. Интересно то, что ни один из элементов питания не может быть заменён на другой – они все нужны, пусть даже некоторые из них в совсем мизерных количествах. И практически каждый из них влияет на фотосинтез – важнейший процесс получения органического вещества из неорганического под воздействием света. В основе фотосинтеза лежит специфическое вещество растений – хлорофилл, и как раз при недостатке любого из элементов питания сразу же нарушается синтез этого уникального зелёного вещества! Этот дефицит сразу же проявляется в виде хлорозов (нарушении естественной окраски листьев) и задержек роста.
Закон «минимума Либиха» гласит, что состояние растения зависит от того фактора, который находится в минимуме. Его ещё называют законом «бочки Либиха», потому что это хорошо отражает суть явления: сколько бы не было длинных досок в этой бочке, достаточно лишь одной короткой, чтобы вода начала вытекать из неё. Наша с вами задача – не допустить дефицита питания для нашей рассады и комнатных растений, поскольку каждый из элементов важен. В органо-минеральном удобрении Богатый Дом подобраны все необходимые для растений питательные элементы с учётом их физиологии.
Его ещё называют законом «бочки Либиха», потому что это хорошо отражает суть явления: сколько бы не было длинных досок в этой бочке, достаточно лишь одной короткой, чтобы вода начала вытекать из неё. Наша с вами задача – не допустить дефицита питания для нашей рассады и комнатных растений, поскольку каждый из элементов важен. В органо-минеральном удобрении Богатый Дом подобраны все необходимые для растений питательные элементы с учётом их физиологии.
Азот нужен растениям для синтеза большинства органических соединений, к примеру, белка – основы жизни. Только при наличии азота в форме нитратов и аммония растения могут выстраивать свой организм. Азот усиливает рост, цветение и плодоношение, улучшает внешний вид растений.
Фосфор в растениях играет очень важную роль в обмене веществ, так как входит в состав большинства органических соединений, таких как ДНК, витамины, фосфолипиды и многих других. С помощью фосфора запасается и расходуется энергия, строятся клеточные мембраны и синтезируются многие вещества в растениях. Усиленное обеспечение растений фосфором ускоряет их развитие и позволяет получать более ранний урожай, одновременно улучшается и его качество.
Усиленное обеспечение растений фосфором ускоряет их развитие и позволяет получать более ранний урожай, одновременно улучшается и его качество.
Калий в растениях отвечает за оводнённость тканей, за всасывающую силу корней. В результате растения, обеспеченные калием, становятся устойчивыми к избытку и к недостаточности влаги, повышенным и пониженным температурам. Калий максимально расходуется растениями в период наибольшего развития надземной массы, то есть, чем больше растение становится – тем больше ему требуется калия.
Бор отвечает за формирование цветков, завязей и полноценных плодов, увеличивает процент завязывания плодов. Играет важную роль в делении клеток и синтезе белков, и является необходимым компонентом клеточной оболочки.
Медь повышает устойчивость растений ко всем неблагоприятным факторам, усиливает интенсивность дыхания, обеспечивает накопление азота, а также нужна растениям для образования хлорофилла, белков и углеводов.
Цинк в растениях входит в состав ферментов, участвует в дыхании, белковом и углеводном обмене, отвечает за образование важной аминокислоты триптофана, повышает содержание фитогормонов, влияющих на накопление биомассы растений. Цинк необходим для нормального развития яйцеклетки и зародыша. Он повышает засухо-, жаро- и холодостойкость растений.
Марганец в растении активирует более 35 ферментов, участвует в фотосинтезе и синтезе витаминов С, В, Е, способствует увеличению содержания сахаров и их оттоку из листьев, ускоряет рост растений и созревание семян.
Кобальт в растениях отвечает за активную симбиотическую азотфиксацию, повышает общее содержание воды в растениях, особенно в засуху. Входит в состав витамина В12, который синтезируется бактериями, в том числе в кишечнике травоядных животных и человека. В связи с этим корма для животных и пища человека должна содержать кобальт.
Молибден в растении играет большую роль в азотном обмене и синтезе белковых веществ, способствует усвоению азота, растворенного в воде, фиксации азота, участвует в синтезе ряда веществ. Под влиянием молибдена в растениях увеличивается содержание углеводов, каротина и аскорбиновой кислоты, белковых веществ, хлорофилла и повышается интенсивность фотосинтеза.
Под влиянием молибдена в растениях увеличивается содержание углеводов, каротина и аскорбиновой кислоты, белковых веществ, хлорофилла и повышается интенсивность фотосинтеза.
Сера нужна растениям для строительства белков, потому что входит в состав серосодержащих аминокислот, регулирует окислительно-восстановительные процессы и участвует в синтезе витаминов и хлорофилла. Способствует усиленному развитию корней растений и симбиотических клубеньковых бактерий, усваивающих атмосферный азот и улучшающих почвенное плодородие.
Литий положительно влияет на развитие корневой системы растений, улучшает транспорт калия, усиливает фотохимическую активность хлоропластов в листьях, влияет на водно-солевой обмен.
Селен в растительном организме влияет на устойчивость к разного рода стрессам: воздействия ядохимикатов, жары, засоления и т.д.
Хром в небольших концентрациях стимулирует активность ряда ферментов, повышает содержание хлорофилла и продуктивность фотосинтеза в листьях.
Никель у растений входит в состав фермента уреазы, который разлагает мочевину до аммиака и углекислого газа, а также активирует ряд ферментов, участвует в перемещении азота и обеспечении им растительных тканей.
Гуми – природная субстанция, по праву носящая звание эликсира роста и плодородия. Присутствуя в препаратах, это сложное органическое вещество придаёт им замечательные свойства. Всё дело в том, что молекулы гуматов – основы Гуми – имеют строение, позволяющее словно клешнями захватывать элементы питания и проводить их в таком состоянии в виде хелатов через клеточные мембраны растений. Химическое родство гуматов и растительной клетки не случайно – их получают из бурого угля. Гуматы лежат в основе гумуса, так называемой «души почвы», дома для всех живых обитателей почвы и «хлеба для растений». Ликвидируется «усталость» почвы, повышает в ней количество полезных микроорганизмов, улучшается её механический состав. Гуми повышает устойчивость к засухе и урожайность культурных растений, так как гуминовые вещества, улучшая физико-химические свойства почвы, способствуют увеличению корневой системы в 1,5–2,5 раза, что позволяет корням проникать в более глубокие слои почвы и потреблять имеющуюся там влагу, в результате чего растения меньше испытывают потребность в поливах.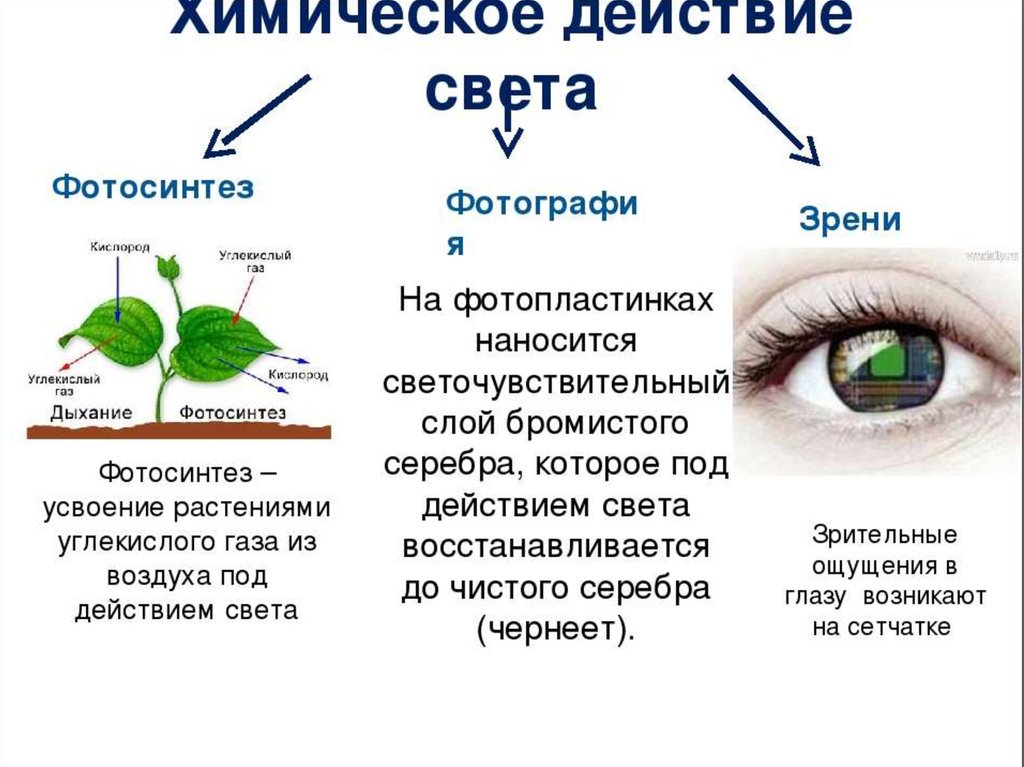 Важно, что Гуми переводит тяжелые металлы, остатки химических загрязнителей и радионуклиды в недоступную для растений форму!
Важно, что Гуми переводит тяжелые металлы, остатки химических загрязнителей и радионуклиды в недоступную для растений форму!
Фитоспорин в Богатом Доме содержится в профилактической дозе, но даже в таких количествах полезные фитобактерии выполняют свою защитную роль – не дают патогенным микроорганизмам развиться и размножиться. Кроме того, они ещё и заводы по производству биологически активных веществ, которые способны стимулировать рост и развитие растений. А, учитывая, что фитобактерии свободно циркулируют по всему растению, можно представить, как быстро достигают биологически активные вещества своих мест назначения.
/фото ГУМИ+Фитоспорин=Богатый Дом/
Всё вышеперечисленное свидетельствует о том, что Богатый Дом недаром носит своё название – такого богатого состава вам не найти ни в одном комплексном удобрении! Удобрительный полив биораствором этого препарата проводим регулярно 1 раз в 2 недели, растворяя всего 1 столовую ложку его в литре чистой воды.
Выращивая перец в домашних условиях с использованием Богатого Дома, я констатировал увеличение урожайности в 2 раза! Урожай получился сладким, сочным и, судя по всему, витаминным. Растения с Богатым Домом жили богато и поэтому поделились с нами своим богатством.
Роль калия в жизни растений — Fitofert Россия
Калий наряду с азотом и фосфором относятся к главным элементам питания растений. Этот важный компонент помогает регулировать водный баланс растений через корни (осмотический градиент) и функционирование устьиц листа. Калий способствует накоплению крахмала и сахара в плодах, увеличивает сопротивляемость растений грибковым и микробным заболеваниям и повреждению насекомыми, и играет важную роль в десятках метаболических реакций, активизируя, по меньшей мере, 60 различных ферментов, участвующих в росте растений, фотосинтезе, в каждом из этапов синтеза белка и обмене веществ в целом. Данный элемент противодействует соляному стрессу от ионов натрия, т.к. калий конкурирует с ним за поглощение корнями.
Данный элемент противодействует соляному стрессу от ионов натрия, т.к. калий конкурирует с ним за поглощение корнями.
Адекватные уровни калия повышают производство и транспортировку углеводов в растении. Серьезный дефицит калия тормозит передачу сахаров внутри растения, приводя к аккумуляции крахмала в нижних листьях. Калий также играет важную роль в повышении устойчивости растений к низким температурам, засолению, засухе и болезням. Поэтому растению необходимо подавать достаточное количество калия для удовлетворения потребностей во всех его частях.
Внесение достаточного количества калия, который поддерживает листву в хорошем состоянии по мере роста растения положительным образом влияет на урожай и на содержание растворимых веществ в плодах (больше сахара) на момент сбора урожая. Более 50% всего потребленного объема калия растением в конечном итоге оказывается в плодах (Таблица 1). Действие, которое оказывает калий на синтез протеинов, усиливает преобразование потребленного нитрата в белки, что способствует эффективному усвоению применяемых азотных удобрений.
Таблица 1. Аккумуляция питательных элементов в сухом веществе различных частей растения перца в % от выноса элементов.
Части растения | Содержание элементов в % от сухого вещества | ||||
N | P | K | Ca | Mg | |
| Ликвидный урожай | 50 | 60 | 50 | 15 | 25 |
| Завязь плодов | 7 | 9 | 7 | 3 | 5 |
| Всего генеративные части растения | 57 | 69 | 57 | 18 | 30 |
| Листва | 25 | 17 | 21 | 60 | 45 |
Стебель | 13 | 10 | 18 | 17 | 21 |
Корни | 5 | 4 | 4 | 5 | 4 |
Всего вегетативные части растения | 43 | 31 | 43 | 82 | 70 |
ВСЕГО по растению | 100 | 100 | 100 | 100 | 100 |
Калий это катион, который вовлечен в поддержание осмотического потенциала растений (клеточный тургор), одним из следствий этого является движения устьиц — отверстий, которые позволяют растениям обмениваться газом и водой с атмосферой. Это позволяет растениям регулировать необходимый водный баланс в условиях стресса, таких как высокое содержание солей и дефицит воды. Действительно, растения с высоким содержанием калия обычно более эффективно используют воду, то есть потребляют сравнительно меньше воды для производства того же количества биомассы, чем растения для которых характерен дефицит калия.
Это позволяет растениям регулировать необходимый водный баланс в условиях стресса, таких как высокое содержание солей и дефицит воды. Действительно, растения с высоким содержанием калия обычно более эффективно используют воду, то есть потребляют сравнительно меньше воды для производства того же количества биомассы, чем растения для которых характерен дефицит калия.
Кроме того калий оказывает влияние на процесс созревания плодов, а именно на синтез пигмента ликопина, который отвечает за красный цвет плодов.
Рисунок 1. Калий усиливает транспорт и хранение продуктов фотосинтеза от листьев к плодам, как показано на рисунке на примере томата.
Калий в почве
Калий, находящийся в почве с точки зрения доступности для растения, можно классифицировать по следующим категориям:
- Недоступный калий
- Связанный частичками почвы или медленно доступный калий
- Обменный калий или готовый к поглощению калий
- Калий в почвенном растворе
Основа вышеназванной классификации – это его доступность для растений. В зависимости от типа почвы и условий окружающей среды, доступность калия может изменяться.
В зависимости от типа почвы и условий окружающей среды, доступность калия может изменяться.
Недоступный калий – находится в виде кристаллов в глиняных жилках почвы, минералов, слюды, которые являются частями почвенной структуры. Растения не могут потреблять данный вид калия в данной нерастворимой форме. Однако, со временем, данные минералы распадаются, и небольшое количество калия переходит в почвенный раствор.
Связанный частичками почвы калий – данный вид калия медленно высвобождается в течение сезона выращивания. Глиняные минералы почвы имеют тенденцию фиксировать калий. В процессе переувлажнения и пересыхания почвы, калий попадает в ловушку между частичками глины (глиняные минералы имеют слоистую структуру). Как только почва увлажняется, некоторые из ионов калия высвобождаются в почвенный раствор. Медленно высвобождаемый калий обычно не подлежит измерению при проведении стандартного анализа почвы.
Обменный калий – доступный для растения калий, который растение может потреблять сразу. Данная фракция калия находится на поверхности частичек глины и органических веществ в почве. Он находится в равновесии с почвенным раствором и легко высвобождается, когда растения поглощают калий из почвенного раствора. Обменный калий измеряется при проведении большинства анализах почвы.
Данная фракция калия находится на поверхности частичек глины и органических веществ в почве. Он находится в равновесии с почвенным раствором и легко высвобождается, когда растения поглощают калий из почвенного раствора. Обменный калий измеряется при проведении большинства анализах почвы.
Калий в почвенном растворе – растворенный калий в почвенном растворе и готовый к абсорбции растением. Это самый маленький резервуар его потребления. Поэтому его анализ в почвенном растворе не дает адекватную картину нахождения данного элемента в почве, доступного для растений.
Фракции калия в почве
Факторы, влияющие на доступность калия в почве.
На доступность калия для растений в почве влияют несколько факторов:
- Уровень кислорода – кислород необходим для надлежащего функционирования корневой системы, включая поглощение калия.
- Влажность – чем больше влаги в почве, тем лучше калий усваивается растениями.
- Обработка почвы — исследование показало, что регулярно обрабатываемая почва позволяет лучше поглощать калий.

- Температура почвы – 15-27°С являются идеальными условиями активности корневой системы и большинства физиологических процессов в растении. Чем ниже температуры, тем ниже абсорбция.

Все основные катионы: кальций, магний, калий и натрий конкурируют друг с другом за поглощения клетками корней. Поэтому вносить их необходимо в сбалансированных пропорциях. Например, внесение чрезмерных доз извести может привести к формированию дефицита магния, перенасыщение калием также приводит к дефициту магния, либо замещает кальций в растениях, создавая тем самым множество проблем и т.д. Чрезмерный натрий замещает калий, что приводит к ряду проблем для растения. Правильное соотношение калия и магния является пропорция 4 :1.
Рисунок 2: Доступность калия через корни растения в зависимости от рН почвенного раствора
Исследования показали, что увеличения нормы содержания калия в период налива и созревания плодов приводит к следующим эффектам:
- Увеличение количество плодов и их веса
- Утолщение стенок плодов
- Улучшение качественных характеристик плодов
Калий увеличивает сопротивляемость растений к заболеваниям.
Калий оказывает хорошо известный эффект повышения устойчивости растений к патогенам и снижения воздействия инфекции:
- Калий снижает эффект бактериоза (пятнистость листьев), вызываемый Pseudomonas lachrymans.
- У партенокарпических тепличных огурцов за счет применения дополнительных доз калия на 27% — 33% снижается серая плесень плодов, вызываемая Botrytis cinerea.
- Листовые обработки солями калия помогают контролировать мучнистую росу, вызываемую Sphaerotheca fuliigiena (Фото 2).
Листовые подкормки нитратом калия помогают растению огурца вырабатывать индивидуальную сопротивляемость к заболеванию.
Фото 2 A: Сверху: лист огурца, инфицированный мучнистой росой.
Фото 2 B: Лист с соседнего растения, обработанного удобрением с содержанием калия.
Дефицит калия
Первичные симптомы дефицита калия на листьях проявляется в виде их окрашивания в темно-зеленый или голубоватый цвет. Листья приобретают чуть блестящую поверхность. Между прожилок листа проявляются пятна ярко-зеленого цвета. У больших листьев края начинают закручиваться вниз в тонкие листочки. На них появляются морщины, а верхняя (адаксиальная) поверхность листа становится коричневой, тогда как обратная сторона листа (абаксиальная) имеет россыпь коричневых точек, похожих на симптомы ранней гнили.
Листья приобретают чуть блестящую поверхность. Между прожилок листа проявляются пятна ярко-зеленого цвета. У больших листьев края начинают закручиваться вниз в тонкие листочки. На них появляются морщины, а верхняя (адаксиальная) поверхность листа становится коричневой, тогда как обратная сторона листа (абаксиальная) имеет россыпь коричневых точек, похожих на симптомы ранней гнили.
Фото 3. Пример дефицита калия на ботве картофеля.
Растения картофеля с дефицитом калия растут медленнее, для них характерна более мелкая листва с неравномерной ребристой поверхностью (Фото 3). Растение приобретает темно-зеленый цвет, чуть позже кромка листа становится бронзовой и желтоватой и данный хлороз распространяется почти на всю поверхность листа.
На землянике первые симптомы дефицита калия проявляются на верхних жилках старых (нижних) листьев. Кончики зубцов краснеют, повреждение постепенно прогрессирует внутрь листа между прожилок до тех пор, пока не затрагивается большая часть листовой пластины.
Фото 4. Дефицит калия на листьях земляники с усугублением симптомов по мере старения листа.
Фото 5. Типичное проявление дефицита калия на листьях огурца.
Образование коричневых пятен (некрозов) связано с нарушением азотного обмена и образованием в тканях яда разложения — путресцина. Дефицит калия также влияет на качество плодов. Плоды деформируются, могут плохо окрашиваться, становятся рыхлыми и безвкусными.
Калий является мобильным элементом в растении и в условиях недостатка движется только к более молодым листьям. Хотя рост растений, испытывающих дефицит калия, не может быть серьезно нарушен, однако урожайность и качество плодов значительно снижаются. Плод не расширяется и не вытягивается в полной мере, а его кончик разбухает, что также может указывать на симптом водного стресса.
Фото 6. Деформация плода из-за набухания кончика.
Решение проблемы
- В средние или тяжелые почвы, где движение калия затруднено, перед посадкой необходимо внести нитрат калия. Определение необходимой нормы внесения должно основываться на анализе почвы.
- В песчаных почвах, где калийные удобрения продвигаются быстрее, внесение водорастворимых макроудобрений в почву, как правило, помогают быстрой коррекции дефицита калия.
- Дефицит калия может быть быстро откорректирован через систему капельного полива (фертигацию).
- Эффективным методом также можно считать листовые подкормки калиевыми препаратами в течение короткого периода времени.
- Для культур, выращиваемых на гидропонике, используют питательный раствор, содержащий 150-200 мг/л K.
 Определение необходимой нормы внесения должно основываться на анализе почвы.
Определение необходимой нормы внесения должно основываться на анализе почвы.Избыток калия
Растения из доступных ему питательных веществ предпочтительно поглощает ионы калия. При чрезмерном внесении калия резко сокращается поглощение, прежде всего магния и кальция. Поэтому на почвах, переудобренных калием, плоды часто страдают от дефицита кальция, а на листьях возникают признаки недостатка магния. Когда калий для растений находится в избытке, в почве затрудняется также и поглощение растением микроэлементов.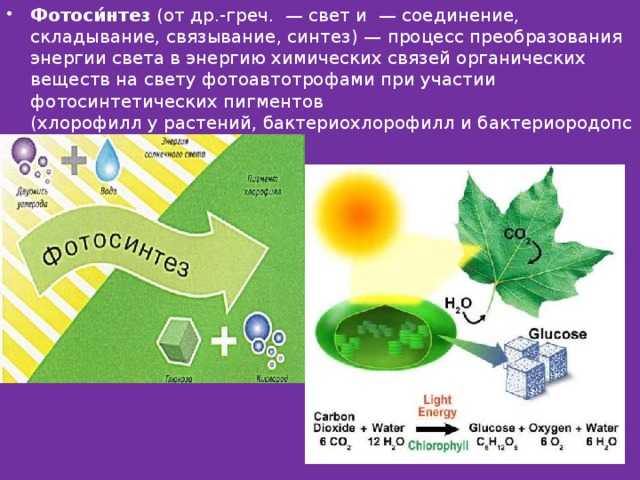
Такие физиологические нарушения, как побурение мякоти, побурение сердечка и загнивание чечевичек, а также преждевременное опадение листьев и др. часто считают следствием чрезмерного удобрения калием.
Особенно опасно излишнее количество калия на фоне недостатка азота и фосфора.
Калийные удобрения лучше вносить несколько раз малыми дозами, чем один раз в большой концентрации. Кроме того, следует знать, что калий действует на растение лучше, если удобрение внесено во влажную почву при прохладной погоде.
Определить потребность калия для растений можно в известной степени на основании данных почвенного анализа. Примерные дозы калийных удобрений при недостаточном содержании калия в почве равны 150—200 кг/га К2О, при нормальном — 100—150 кг/га К2О; при избыточном содержании калий не вносят.
Калийные удобрения
Основными источниками калия являются: нитрат калия, сульфат калия, сульфат калия-магния и хлорид калия. Самый быстрый источник в доступной для растения форме это калиевая селитра. Данное удобрение является идеальным удобрением на стадии налива плодов. Из-за своей хорошей мобильности он может быть потреблен растением через час после внесения в почву. Благодаря своему взаимодействию с ионами (нитратами) он также важен на данной стадии развития, он требует минимального количества влаги в почве для растворения и доступности растения. Другие источники калия имеют преимущество из-за более низкой стоимости, однако они имеют и свои недостатки.
Например, сульфат калия имеет более низкую растворимость, чем нитрат калия. Второй пример: хлорид калия поставляет большое количество хлоридов, которые в больших дозах являются токсичными для растений. Учитывая негативное влияние хлора на культуры, хлористый калий вносят в грунт только на зиму, перед вспашкой.
Третий вариант: калимагнезия (сульфат калия-магния) слишком медленно преобразуется в доступную форму. Из-за перечисленных особенностей рекомендуется использовать данные источники калия в небольших количествах или только на начальных этапах развития растения, чтобы обеспечить переход удобрений в доступную форму и избежать накопления токсичности из-за хлоридов. Кроме того почва должна получить достаточное количество осадков для вымывания хлоридов, которые могут навредить растению.
Кроме того почва должна получить достаточное количество осадков для вымывания хлоридов, которые могут навредить растению.
Один из основных недостатков наличия хлоридов в почве это их конкуренция с ионами нитратов, фосфатов и сульфатов за вход в растение; чем больше хлоридов проникает в растение, тем меньше других анионов попадет в растение. Хлориды также повышают общий уровень солей в почве и поэтому повышают уровень ЕС. При достижении определенного уровня засоленности, уровень потребление растением воды снижается. Данный феномен может привести к повышению стресса из-за снижения влажности почвы в результате чего размер плодов уменьшаются, а урожайность, соответственно, снижается.
Таблица 2. Характеристики калийных удобрений.
| Общее название | Формула | Характеристики |
| Калиевая селитра | KNO3 | Является идеальным калийным удобрением на всех этапах роста, также является частичным источником нитратного азота для растения. Имеет высокую растворимость 320 г/л при 20°С. Имеет высокую растворимость 320 г/л при 20°С. |
| Сульфат калия | K2SO4 | Является идеальным калийным удобрением на поздней стадии роста, когда растению не требуется азот. Сульфат калия имеет ограниченную растворимость около 6% (при смешивании с другими удобрениями). |
| Бикарбонат калия | К(НСО3)2 | Используется в основном как регулятор рН для его повышения. |
| Хлорид калия | KCl | Хлориды не рекомендуется широко использоваться для культур, т.к. они чувствительны к высокому содержанию солей в корневой зоне. Использование хлористых удобрений приводит к конкуренции в корневой зоне между анионами (NO3—, h3PO4—,SO42-), что в конечном итоге приводит к нарушению баланса питательных веществ.
|
Еще раз перечислим основные функции калия и эффект его внесения.
При достаточном поступлении калия в растение:
- Процессы окисления в клетках идут интенсивнее
- Усиливается клеточный обмен
- Растение легче переносит недостаток влаги
- Фотосинтез ускоряется
- Возрастает ферментативная активность
- Легче проходит обмен белков и углеводов
- Растения быстрее адаптируются к отрицательным температурам
- Образуется больше органических кислот
- Повышается сопротивляемость к патогенным факторам
При нехватке калия:
- Из простых углеводов не синтезируются сложные
- Образование белка в клетках прекращается.
- Происходит задержка в развитии репродуктивных органов.
- Стебель становится слабым.

Все вышеперечисленные факторы прямо или косвенно влияют на формирование плодов и их качества, а значит и на вашу прибыль!
Линейка комплексных водорастворимых NPK-удобрений ФИТОФЕРТ ЭНЕРДЖИ содержит сбалансированный состав всех основных элементов питания в зависимости от потребностей растений на каждом этапе его развития, способствуя их эффективному росту и развитию!
ФИТОФЕРТ ЭНЕРДЖИ 5-55-10 СТАРТ– на начальном этапе растение не нуждается в больших количествах калия, поэтому в данном удобрении он представлен в виде монофосфата калия. Соотношение активных K и P в препарате оптимально для максимального плодоношения, а также повышения стойкости растений к болезням, вредителям и заморозкам.
ФИТОФЕРТ ЭНЕРДЖИ 20-20-20 БАЛАНС – данное удобрение содержит сбалансированный состав по всем основным элементам, калий в нем представлен в виде быстродоступного нитрата калия и монофосфата калия.
ФИТОФЕРТ ЭНЕРДЖИ 15-5-33 АКТИВ – на этапе налива плодов одним из основных элементов является калий, в состав данного удобрения входят 3 видов калийных удобрений, 50% нитрата калия, как самого быстрого и легкодоступного источника калия, 22% нитрата аммония, позволяющий получить калий в более отсроченное время, обеспечивая его непрерывную подачу, и монофосафата калия.
ФИТОФЕРТ ЭНЕРДЖИ 0-15-45 ФИНИШ – данное удобрение применяется на финальных этапах формирования подов – окрашивания и созревания, поэту нитратная форма калийных удобрений в нем исключена. В данном удобрении калий представлен в виде сульфата, которые способствуют созреванию плодов и монофосфата калия, когда растениям нужна дополнительная энергия, которую обеспечивает фосфор.
Применение калия повышает фотосинтез и биосинтез сорбитола, а также ускоряет акклиматизацию подорожника обыкновенного (Plantago major L.) к холоду
1. Hawkesford M., Horst W., Kichey T., Lambers H.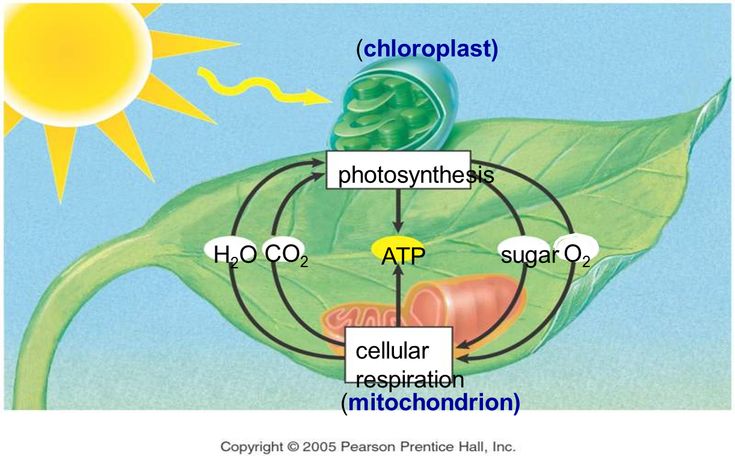 , Schjoerring J., Møller I.S., White P. Глава 6 — Функции макронутриентов. В: Маршнер П., редактор. Минеральное питание высших растений Маршнера. 3-е изд. Академическая пресса; Сан-Диего, Калифорния, США: 2012. стр. 135–189. [Google Scholar]
, Schjoerring J., Møller I.S., White P. Глава 6 — Функции макронутриентов. В: Маршнер П., редактор. Минеральное питание высших растений Маршнера. 3-е изд. Академическая пресса; Сан-Диего, Калифорния, США: 2012. стр. 135–189. [Google Scholar]
2. Рем Г., Шмитт М. Калий для растениеводства: управление питательными веществами. Расширение Университета Миннесоты; Миннеаполис, Миннесота, США: 2002. [Google Scholar]
3. Ragel P., Raddatz N., Leidi E.O., Quintero F.J., Pardo J.M. Регулирование питания K+ в растениях. Фронт. Растениевод. 2019;10:281. doi: 10.3389/fpls.2019.00281. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
4. Roelfsema M.R.G., Hedrich R. В свете открывания устьиц: новый взгляд на «Уотергейт» New Phytol. 2005; 167: 665–691. doi: 10.1111/j.1469-8137.2005.01460.x. [PubMed] [CrossRef] [Google Scholar]
5. Cochrane T.T., Cochrane T.A. Жизненно важная роль калия в осмотическом механизме модуляции апертуры устьиц и его связь с дефицитом калия. Сигнал завода. Поведение 2009 г.;4:240–243. doi: 10.4161/psb.4.3.7955. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Сигнал завода. Поведение 2009 г.;4:240–243. doi: 10.4161/psb.4.3.7955. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Канаи С., Могаиб Р.Э., Эль-Шеми Х.А., Паниграхи Р., Мохапатра П.К., Ито Дж., Нгуен Н.Т., Санеока Х., Фуджита K. Дефицит калия влияет на водный статус и скорость фотосинтеза вегетативного поглотителя томатов в теплицах до того, как он повлияет на активность источника. Растениевод. 2011; 180:368–374. doi: 10.1016/j.plantsci.2010.10.011. [PubMed] [CrossRef] [Google Scholar]
7. Hosseini S.A., Hajirezaei M.R., Seiler C., Sreenivasulu N., von Wirén N. Потенциальная роль калия флагового листа в придании устойчивости ячменю к старению листьев, вызванному засухой. . Фронт. Растениевод. 2016;7:206. дои: 10.3389/fpls.2016.00206. [Статья PMC free] [PubMed] [CrossRef] [Google Scholar]
8. Дрекслер Н., Чжэн Ю., Бонер А., Нобманн Б., фон Вирен Н., Кунце Р., Рауш С. Зависимость от нитратов контроль гомеостаза побегов K с помощью члена семейства транспортеров нитратов / переносчиков пептидов NPF7. 3/НРТ1. 5 и звездчатый K+ выпрямитель SKOR в арабидопсисе. Завод Физиол. 2015;169:2832–2847. [PMC free article] [PubMed] [Google Scholar]
3/НРТ1. 5 и звездчатый K+ выпрямитель SKOR в арабидопсисе. Завод Физиол. 2015;169:2832–2847. [PMC free article] [PubMed] [Google Scholar]
9. Etienne P., Diquelou S., Prudent M., Salon C., Maillard A., Ourry A. Запас макро- и микроэлементов в растениях и их ремобилизация при столкновении дефицит: случай засухи. Сельское хозяйство. 2018;8:14. дои: 10.3390/сельское хозяйство8010014. [CrossRef] [Google Scholar]
10. Уайт П. Дж. Глава 3 — Перенос на большие расстояния в ксилеме и флоэме. В: Маршнер П., редактор. Минеральное питание высших растений Маршнера. 3-е изд. Академическая пресса; Сан-Диего, Калифорния, США: 2012. стр. 49–70. [Google Scholar]
11. Поммеррениг Б., Эггерт К., Бинерт Г.П. Влияние дефицита бора на профили сахара, ионома и фитогормонов сосудистых и несосудистых тканей листьев подорожника обыкновенного ( Plantago major Л.) Междунар. Дж. Мол. науч. 2019;20:3882. doi: 10.3390/ijms20163882. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Ruiz-Medrano R., Xoconostle-Cázares B., Lucas WJ. Флоэма как канал межорганной коммуникации. Курс. мнение биол. растений 2001; 4: 202–209. doi: 10.1016/S1369-5266(00)00162-X. [PubMed] [CrossRef] [Google Scholar]
Ruiz-Medrano R., Xoconostle-Cázares B., Lucas WJ. Флоэма как канал межорганной коммуникации. Курс. мнение биол. растений 2001; 4: 202–209. doi: 10.1016/S1369-5266(00)00162-X. [PubMed] [CrossRef] [Google Scholar]
13. Wippel K., Sauer N. Arabidopsis SUC1 загружает флоэму у мутантов suc2 при экспрессии с промотора SUC2. Дж. Эксп. Бот. 2011;63:669–679. doi: 10.1093/jxb/err255. [Статья PMC free] [PubMed] [CrossRef] [Google Scholar]
14. Чакмак И., Хенгелер С., Маршнер Х. Изменения экспорта сахарозы флоэмой в листьях в ответ на дефицит фосфора, калия и магния у бобовых растений . Дж. Эксп. Бот. 1994;45:1251–1257. doi: 10.1093/jxb/45.9.1251. [CrossRef] [Google Scholar]
15. Koch M., Busse M., Naumann M., Jakli B., Smit I., Cakmak I., Hermans C., Pawelzik E. Дифференциальные эффекты разнообразного питания калием и магнием на продукцию и распределение фотоассимилятов в растениях картофеля. Физиол. Завод. 2019;166:921–935. doi: 10.1111/ppl.12846. [PubMed] [CrossRef] [Google Scholar]
16. Gajdanowicz P., Michard E., Sandmann M., Rocha M., Corrêa L.G.G., Ramirez-Aguilar S.J., Gomez-Porras J.L., Gonzalez W., Thibaud J. -Б., Ван Донген Дж.Т. Градиенты калия (К+) служат мобильным источником энергии в сосудистых тканях растений. проц. Натл. акад. науч. США. 2011; 108: 864–869. doi: 10.1073/pnas.1009777108. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Gajdanowicz P., Michard E., Sandmann M., Rocha M., Corrêa L.G.G., Ramirez-Aguilar S.J., Gomez-Porras J.L., Gonzalez W., Thibaud J. -Б., Ван Донген Дж.Т. Градиенты калия (К+) служат мобильным источником энергии в сосудистых тканях растений. проц. Натл. акад. науч. США. 2011; 108: 864–869. doi: 10.1073/pnas.1009777108. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
17. Дрейер И., Гомес-Поррас Дж.Л., Ридельсбергер Дж. Калиевая батарея: мобильный источник энергии для транспортных процессов в сосудистых тканях растений. Новый Фитол. 2017;216:1049–1053. doi: 10.1111/nph.14667. [PubMed] [CrossRef] [Google Scholar]
18. Seki M., Umezawa T., Urano K., Shinozaki K. Регуляторные метаболические сети в реакциях на засуху и стресс. Курс. мнение биол. растений 2007; 10: 296–302. doi: 10.1016/j.pbi.2007.04.014. [PubMed] [CrossRef] [Google Scholar]
19. Pommerrenig B., Cvetkovic J., Trentmann O., Klemens P.A.W., Neuhaus H.E., Ludewig F. In Concert: Организованные изменения в гомеостазе углеводов имеют решающее значение для устойчивости растений к абиотическим стрессам. . Физиология клеток растений. 2018;59: 1290–1299. doi: 10.1093/pcp/pcy037. [PubMed] [CrossRef] [Google Scholar]
. Физиология клеток растений. 2018;59: 1290–1299. doi: 10.1093/pcp/pcy037. [PubMed] [CrossRef] [Google Scholar]
20. Ядав С.К. Механизмы устойчивости растений к холодовому стрессу. Обзор. Агрон. Поддерживать. Дев. 2010;30:515–527. doi: 10.1051/agro/2009050. [CrossRef] [Google Scholar]
21. Strand Å., Hurry V., Gustafsson P., Gardeström P. Развитие листьев Arabidopsis thaliana при низких температурах приводит к подавлению фотосинтеза и экспрессии фотосинтетических генов, несмотря на накопление растворимых углеводов. Завод Ж. 1997;12:605–614. doi: 10.1046/j.1365-313X.1997.00583.x. [PubMed] [CrossRef] [Google Scholar]
22. Zuther E., Büchel K., Hundertmark M., Stitt M., Hincha D.K., Heyer AG. Роль раффинозы в реакции акклиматизации Arabidopsis thaliana на холод. ФЭБС лат. 2004; 576: 169–173. doi: 10.1016/j.febslet.2004.09.006. [PubMed] [CrossRef] [Google Scholar]
23. Пешев Д., Вергаувен Р., Моглиа А., Хидег Э., Ван ден Энде В. На пути к пониманию вакуолярных антиоксидантных механизмов: роль фруктанов? Дж. Эксп. Бот. 2013;64:1025–1038. дои: 10.1093/jxb/ers377. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Эксп. Бот. 2013;64:1025–1038. дои: 10.1093/jxb/ers377. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Бхаттачарья С., Кунду А. Сахара и полиолы сахаров в преодолении экологических стрессов. В: Ройчоудхури А., Трипати Д.К., редакторы. Защитные химические агенты в смягчении абиотического стресса растений: биохимические и молекулярные перспективы. Джон Уайли и сыновья; Хобокен, Нью-Джерси, США: 2020. с. 71. [Google Scholar]
25. Stoop J.M., Williamson J.D., Pharr D.M. Метаболизм маннита в растениях: метод борьбы со стрессом. Тенденции Растениевод. 1996;1:139–144. doi: 10.1016/S1360-1385(96)80048-3. [CrossRef] [Google Scholar]
26. Поммеррениг Б., Папини-Терзи Ф.С., Зауэр Н. Дифференциальная регуляция загрузки сорбита и сахарозы во флоэму подорожника большого в ответ на солевой стресс. Завод Физиол. 2007; 144:1029. doi: 10.1104/стр.106.089151. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
27. Nadwodnik J., Lohaus G. Субклеточные концентрации сахарных спиртов и сахаров в связи с транслокацией флоэмы у Plantago major, Plantago maritima, Prunus persica и Apium гравеоленс. Планта. 2008;227:1079–1089. doi: 10.1007/s00425-007-0682-0. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Планта. 2008;227:1079–1089. doi: 10.1007/s00425-007-0682-0. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
28. Ramsperger-Gleixner M., Geiger D., Hedrich R., Sauer N. Дифференциальная экспрессия генов транспортеров сахарозы и полиолов во время созревания подорожника обыкновенного клетки-спутницы. Завод Физиол. 2004; 134:147–160. doi: 10.1104/стр.103.027136. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
29. Ван М., Чжэн К., Шен К., Го С. Критическая роль калия в реакции растений на стресс. Междунар. Дж. Мол. науч. 2013;14:7370–7390. doi: 10.3390/ijms14047370. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Römheld V., Kirkby E.A. Исследования по калию в сельском хозяйстве: потребности и перспективы. Растительная почва. 2010; 335:155–180. doi: 10.1007/s11104-010-0520-1. [CrossRef] [Google Scholar]
31. Hosseini S.A., Maillard A., Hajirezaei M.R., Ali N., Schwarzenberg A., Jamois F., Yvin J.-C. Индукция переносчиков кремния ячменя HvLsi1 и HvLsi2, повышение концентрации кремния в побегах и регулирование гомеостаза крахмала и АБК в условиях осмотического стресса и сопутствующего дефицита калия. Фронт. Растениевод. 2017;8:1359. doi: 10.3389/fpls.2017.01359. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Фронт. Растениевод. 2017;8:1359. doi: 10.3389/fpls.2017.01359. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Деви Б.С.Р., Ким Ю.Дж., Селви С.К., Гаятри С., Алтанзул К., Парвин С., Ян Д.-У., Ли О.Р., Ли С., Ян Д.-К. Влияние нитрата калия на уровень антиоксидантов и гены вторичных метаболитов при холодовом стрессе у женьшеня обыкновенного. Русь. J. Физиол растений. 2012;59:318–325. doi: 10.1134/S1021443712030041. [CrossRef] [Google Scholar]
33. Pommerrenig B., Junker A., Abreu I., Bieber A., Fuge J., Willner E., Bienert M.D., Altmann T., Bienert G.P. Идентификация рапса ( Brassica napus ) Сорта с высокой устойчивостью к условиям дефицита бора. Фронт. Растениевод. 2018;9:1142. doi: 10.3389/fpls.2018.01142. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Jakobsen S.T. Взаимодействие между элементами питания растений: III. Антагонизм между калием, магнием и кальцием. Акта Агрик. Сканд. B-Plant Почвоведение. 1993; 43:1–5. doi: 10.1080/09064719309410223. [CrossRef] [Google Scholar]
doi: 10.1080/09064719309410223. [CrossRef] [Google Scholar]
35. Diem B., Godbold D. Антагонизм калия, кальция и магния в клонах Populus trichocarpa. Растительная почва. 1993;155:411–414. doi: 10.1007/BF00025070. [CrossRef] [Google Scholar]
36. Тума Ю., Скалицкий М., Тумова Л., Блахова П., Росулькова М. Содержание калия, магния и кальция в отдельных частях растения Phaseolus vulgaris L. в зависимости от калия и питание магнием. Почвенная среда растений. 2004; 50:18–26. doi: 10.17221/3637-PSE. [CrossRef] [Google Scholar]
37. Менгель К., Киркби Э. Принципы питания растений. Международный калийный институт; Берн, Швейцария: 1982. [Google Scholar]
38. Дибб Д., Томпсон В. мл. Взаимодействие калия с другими питательными веществами. Калий Агрик. 1985; 1: 515–533. [Google Scholar]
39. Ян Дж., Чжан Дж., Ли К., Чжан З., Ма Ф., Ли М. Реакция метаболизма сахаров в листьях яблони на кратковременный стресс от засухи. Завод Физиол. Биохим. 2019; 141:164–171. doi: 10.1016/j.plaphy.2019.05.025. [PubMed] [CrossRef] [Google Scholar]
doi: 10.1016/j.plaphy.2019.05.025. [PubMed] [CrossRef] [Google Scholar]
40. Ву Д.П., Мартинс Родригес К., Юнг Б., Мейснер Г., Клеменс П.А., Хольтгрёв Д., Фюртауэр Л., Нэгеле Т., Ниберл П., Поммеррениг B. Вакуолярный гомеостаз сахарозы имеет решающее значение для развития растений, свойств семян и ночного выживания арабидопсиса. Дж. Эксп. Бот. 2020;71:4930–4943. doi: 10.1093/jxb/eraa205. [PubMed] [CrossRef] [Google Scholar]
41. Родригес С.М., Мюдсам С., Келлер И., Цирер В., Чарнецкий О., Коррал Дж.М., Рейнхардт Ф., Ниберл П., Фидлер-Вихерс К., Зоммер Ф.К. Яровизация изменяет идентичность стока и источника и обращает транслокацию флоэмы из стержневых корней в побеги в растительной клетке сахарной свеклы ( Beta vulgaris ). 2020 г.: 10.1105/tpc.20.00072. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
42. Германс С., Хаммонд Дж. П., Уайт П. Дж., Вербрюгген Н. Как растения реагируют на нехватку питательных веществ путем распределения биомассы? Тенденции Растениевод. 2006; 11: 610–617. doi: 10.1016/j.tplants.2006.10.007. [PubMed] [CrossRef] [Академия Google]
2006; 11: 610–617. doi: 10.1016/j.tplants.2006.10.007. [PubMed] [CrossRef] [Академия Google]
43. Deeken R., Geiger D., Fromm J., Koroleva O., Ache P., Langenfeld-Heyser R., Sauer N., May S.T., Hedrich R. Потеря калиевого канала AKT2/3 влияет на сахар загружается во флоэму арабидопсиса. Планта. 2002; 216:334–344. doi: 10.1007/s00425-002-0895-1. [PubMed] [CrossRef] [Google Scholar]
44. Conti T.R., Geiger D.R. Калийное питание и транслокация в сахарной свекле. Завод Физиол. 1982; 70: 168–172. doi: 10.1104/стр.70.1.168. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Фюртауэр Л., Вайсманн Дж., Векверт В., Нэгеле Т. Динамика метаболизма растений при акклиматизации к холоду. Междунар. Дж. Мол. науч. 2019;20:5411. doi: 10.3390/ijms20215411. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
46. Cao W., Tibbits T.W. Влияние концентрации калия на рост, газообмен и накопление минералов в картофеле. J. Питательные вещества для растений. 1991; 14: 525–537. doi: 10.1080/01
1991; 14: 525–537. doi: 10.1080/01
47. Джордж М.С., Лу Г., Чжоу В. Генотипические вариации поглощения и эффективности использования калия в батате ( Ipomoea batatas L.) Полевые культуры Res. 2002; 77: 7–15. doi: 10.1016/S0378-4290(02)00043-6. [CrossRef] [Google Scholar]
48. Фарбер А., Янг А.Дж., Рубан А.В., Хортон П., Янс П. Динамика активности ксантофиллового цикла в различных антенных субкомплексах фотосинтетических мембран высших растений (взаимосвязь между зеаксантином конверсия и нефотохимическое тушение флуоресценции) Физиол. растений. 1997; 115:1609–1618. doi: 10.1104/стр.115.4.1609. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Kress E., Jahns P. Динамика диссипации энергии и конверсии ксантофилла у арабидопсиса указывает на косвенную фотозащитную роль зеаксантина в медленно индуцируемых и релаксирующих компонентах нефотохимического тушения энергии возбуждения. Фронт. Растениевод. 2017;8:2094. doi: 10.3389/fpls.2017.02094. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2017;8:2094. doi: 10.3389/fpls.2017.02094. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Асада К. Круговорот воды в хлоропластах: удаление активного кислорода и рассеивание избыточных фотонов. Анну. Преподобный завод биол. 1999;50:601–639. doi: 10.1146/annurev.arplant.50.1.601. [PubMed] [CrossRef] [Google Scholar]
51. Shen C., Li Y., Wang J., Al Shoffe Y., Dong C., Shen Q., Xu Y. Калий влияет на экспрессию ключевых генов, участвующих в Метаболизм сорбита и его ассимиляция в листьях и плодах груши. J. Регулятор роста растений. 2018; 37: 883–895. doi: 10.1007/s00344-018-9783-1. [CrossRef] [Google Scholar]
52. Mengel K., Haeder H.-E. Влияние обеспеченности калием на скорость экссудации флоэмного сока и состав флоэмного сока Ricinus communis. Завод Физиол. 1977;59:282–284. doi: 10.1104/стр.59.2.282. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
53. Benlloch-González M., Romera J., Cristescu S., Harren F., Fournier J.M., Benlloch M. индуцированное закрытие устьиц посредством синтеза этилена в растениях подсолнечника. Дж. Эксп. Бот. 2010;61:1139–1145. doi: 10.1093/jxb/erp379. [PubMed] [CrossRef] [Google Scholar]
индуцированное закрытие устьиц посредством синтеза этилена в растениях подсолнечника. Дж. Эксп. Бот. 2010;61:1139–1145. doi: 10.1093/jxb/erp379. [PubMed] [CrossRef] [Google Scholar]
54. Первез Х., Ашраф М., Махдум М.И. Влияние калийного питания на характеристики газообмена и водные отношения хлопчатника ( Gossypium hirsutum L.) Фотосинтетика. 2004; 42: 251–255. doi: 10.1023/B:PHOT.0000040597.62743.5b. [CrossRef] [Google Scholar]
55. Трентманн О., Мюльхаус Т., Циммер Д., Зоммер Ф.К., Шрода М., Хаферкамп И., Келлер И., Поммеррениг Б., Нойхаус Х.Е. Идентификация белков оболочки хлоропластов, имеющих решающее значение для адаптации к холоду. Завод Физиол. 2020;182:1239–1255. doi: 10.1104/стр.19.00947. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
56. Niittylä T., Messerli G., Trevisan M., Chen J., Smith A.M., Zeeman S.C. Ранее неизвестный переносчик мальтозы, необходимый для деградации крахмала в листья. Наука. 2004; 303: 87–89.. doi: 10.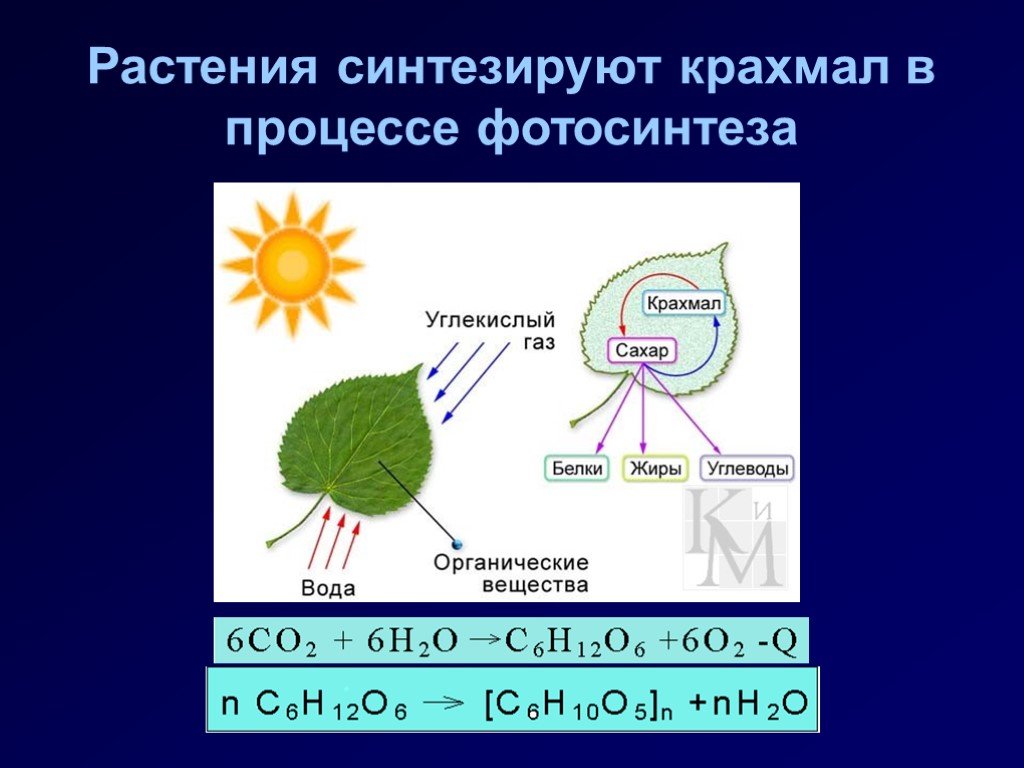 1126/science.1091811. [PubMed] [CrossRef] [Google Scholar]
1126/science.1091811. [PubMed] [CrossRef] [Google Scholar]
57. Klemens P.A.W., Patzke K., Trentmann O., Poschet G., Büttner M., Schulz A., Marten I., Hedrich R., Neuhaus H.E. Сверхэкспрессия протонно-связанного вакуолярного экспортера глюкозы ухудшает устойчивость к замораживанию и прорастание семян. Новый Фитол. 2014; 202:188–197. doi: 10.1111/nph.12642. [PubMed] [CrossRef] [Google Scholar]
58. Huang J., Huang Z., Zhou X., Xia C., Imran M., Wang S., Xu C., Zha M., Liu Y., Чжан С. Тканеспецифическое транскриптомное профилирование подорожника большого дает представление об участии сосудистой сети в реакциях на дефицит фосфатов. Мол. Жене. Геном. 2019;294:159–175. doi: 10.1007/s00438-018-1496-4. [PubMed] [CrossRef] [Google Scholar]
59. Gahrtz M., Stolz J., Sauer N. Специфичный для флоэмы симпортер сахарозы-H+ из Plantago major L. поддерживает модель апопластической загрузки флоэмы. Плант Дж. 1994; 6: 697–706. doi: 10.1046/j.1365-313X.1994.6050697.x. [PubMed] [CrossRef] [Google Scholar]
60. Spitzer M., Wildenhain J., Rappsilber J., Tyers M. BoxPlotR: веб-инструмент для создания коробчатых диаграмм. Нац. Методы. 2014;11:121. doi: 10.1038/nmeth.2811. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Spitzer M., Wildenhain J., Rappsilber J., Tyers M. BoxPlotR: веб-инструмент для создания коробчатых диаграмм. Нац. Методы. 2014;11:121. doi: 10.1038/nmeth.2811. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Калий для растениеводства | Расширение UMN
- Дом
- Растениеводство
- Управление питанием
- Фосфор и калий
- Калий для растениеводства
Калий (K) является важным питательным веществом для роста растений. Он классифицируется как макроэлемент, потому что растения поглощают большое количество калия в течение своего жизненного цикла.
Он классифицируется как макроэлемент, потому что растения поглощают большое количество калия в течение своего жизненного цикла.
Почвы штата Миннесота могут обеспечить некоторое количество калия для выращивания сельскохозяйственных культур, но когда поступления из почвы недостаточно, программа удобрений должна обеспечивать его.
Здесь мы дадим вам общее представление о калии, включая питание растений калием, его реакцию в почве, его функцию в растениях и его роль в эффективном растениеводстве. Кроме того, вы найдете информацию об испытаниях почвы, источниках калия, прогнозировании потребности в калии и эффективном применении калия на ваших полях.
Роль в росте растений
Калий связан с движением воды, питательных веществ и углеводов в тканях растений. Он связан с активацией ферментов в растении, что влияет на производство белка, крахмала и аденозинтрифосфата (АТФ). Производство АТФ может регулировать скорость фотосинтеза.
Калий также помогает регулировать открытие и закрытие устьиц, что регулирует обмен водяного пара, кислорода и углекислого газа. Если K недостаточен или не поступает в достаточном количестве, он останавливает рост растений и снижает урожайность.
Для многолетних культур, таких как люцерна, калий играет важную роль в устойчивости насаждения в течение зимы. Другие роли K включают:
Усиливает рост корней и повышает засухоустойчивость.
Поддерживает тургор; уменьшает потерю воды и увядание.
Участвует в фотосинтезе и формировании пищи.
Уменьшает дыхание, предотвращая потери энергии.
Усиливает транслокацию сахаров и крахмала.
Производит зерно, богатое крахмалом.
Увеличивает содержание белка в растениях.
Создает целлюлозу и уменьшает полегание.
Помогает задерживать болезни растений.

Калий в почвах
Общее содержание калия в почвах часто превышает 20 000 ppm (частей на миллион). В то время как общий запас калия в почве довольно велик, относительно небольшое его количество доступно для роста растений в любой момент времени. Это потому, что почти весь этот K находится в структурном компоненте почвенных минералов и недоступен для роста растений.
Количество K, поставляемого почвами, варьируется из-за больших различий в исходных материалах почвы и влиянии выветривания на эти материалы. Таким образом, потребность в K в программе удобрений варьируется в зависимости от Соединенных Штатов.
Три формы K – недоступные, медленно доступные или фиксированные и легкодоступные или обменные – существуют в равновесии в почвенной системе. Ниже мы опишем эти формы и их взаимосвязь друг с другом. Рисунок 1 также иллюстрирует общую взаимосвязь между этими формами.
Ниже мы опишем эти формы и их взаимосвязь друг с другом. Рисунок 1 также иллюстрирует общую взаимосвязь между этими формами.
Рисунок 1: Обобщенный почвенный цикл K в почве и где K может ежегодно вноситься или удаляться.
Формы калия
|
Поглощение растениями: ключевые факторы
На поглощение калия растениями влияет несколько факторов, включая влажность почвы, аэрацию почвы и уровень кислорода, температуру почвы и систему обработки почвы.
|
Симптомы дефицита калия
Некоторые культуры проявляют характерные симптомы дефицита, когда недостаточное количество калия недоступно для роста и развития. Калий подвижен в растениях и перемещается из нижних листьев в верхние.
Калий подвижен в растениях и перемещается из нижних листьев в верхние.
|
Рисунок 5: Ранние симптомы дефицита калия у картофеля.
Рисунок 6: Поздние симптомы дефицита калия у картофеля.
Прогнозирование потребности в калии
Вы можете отслеживать статус K в почвах с помощью анализа растений и рутинных анализов почвы.
Тестирование почвы является наиболее надежным предиктором потребности программы удобрений в калийных удобрениях. Анализ растений может подтвердить предполагаемый дефицит, на который указывают визуальные симптомы, или регулярно отслеживать эффекты выбранной программы удобрений.
Анализ растений может подтвердить предполагаемый дефицит, на который указывают визуальные симптомы, или регулярно отслеживать эффекты выбранной программы удобрений.
|
Таблица 1: Достаточность калия для основных сельскохозяйственных культур, овощей и фруктов, выращиваемых в Миннесоте
| Урожай | Часть растения | Время | Достаточный диапазон |
|---|---|---|---|
| Люцерна | Топы (новый рост 6 дюймов) | Перед цветением | от 2,0 до 3,5 % К |
| яблоко | Лист из середины текущего побега | 15 июля — 15 августа | от 1,2 до 1,8 % К |
| Черника | Молодой зрелый лист | Первая неделя сбора урожая | от 0,4 до 0,7% К |
| Брокколи | Молодой зрелый лист | Товарная позиция | от 2,0 до 4,0 % К |
| Капуста | Наполовину выросший молодой лист обертки | Головки | от 3,0 до 5,0 % К |
| Морковь | Молодой зрелый лист | Среднерослые | от 2,8 до 4,3 % К |
| Цветная капуста | Молодой зрелый лист | Застегивание пуговиц | от 2,6 до 4,2 % К |
| Кукуруза | Целые вершины | Высота менее 12 дюймов | от 2,5 до 3,5% К |
| Кукуруза | Лист у основания уха | Начальный шелк | от 1,8 до 3,0 % К |
| Съедобная фасоль | Последнее созревшее тройчатое растение | Сцена цветения | от 1,5 до 3,3 % К |
| Виноград | Черешок молодого зрелого листа | Цветение | от 1,5 до 2,0 % К |
| Горох | Недавно созревшая листовка | Первое цветение | от 2,0 до 3,5 % К |
| Картофель | Четвертый лист от кончика | Через 40-50 дней после появления всходов | от 4,0 до 6,0 % К |
| Картофель | Черешок от четвертого листа до верхушки | Через 40-50 дней после появления всходов | от 8,0 до 10,0 % К |
| Малиновый | Лист 18 дюймов от кончика | Первая неделя августа | от 1,1 до 3,0% К |
| Соя | Тройчатые листья | Раннее цветение | от 1,7 до 2,5 % К |
| Яровая пшеница | Целые вершины | Когда голова выходит из ботинка | от 1,5 до 3,0 % К |
| Клубника | Молодой зрелый лист | Середина августа | от 1,1 до 2,5 % К |
| Сахарная кукуруза | Початок | Кисточка для шелка | от 1,8 до 3,0 % К |
| Сахарная свекла | Недавно созревшие листья | Через 50–80 дней после посева | от 2,0 до 6,0 % К |
Источник: Bryson et al. (2014) и Розен и Элиасон (2002). (2014) и Розен и Элиасон (2002). |
Таблица 2: Относительные уровни значений K в тесте почвы для высушенных на воздухе образцов почвы штата Миннесота
| Почвенный калий | Относительный уровень |
|---|---|
| от 0 до 40 частей на миллион (частей на миллион) | Очень низкий |
| от 41 до 80 частей на миллион | Низкий |
| от 81 до 120 частей на миллион | Средний |
| от 121 до 160 частей на миллион | Высокий |
| 161+ частей на миллион | Очень высокая |
Тесты почвы: влияние высыхания образца почвы на результаты
Лаборатории по тестированию почвы обычно высушивают образцы почвы перед анализом. Высыхание почв с высоким содержанием глины может повлиять на количество извлекаемого калия.
Высыхание почв с высоким содержанием глины может повлиять на количество извлекаемого калия.
|
Таблица 3: Общие источники калийных удобрений
| Материал | Химическая формула | Приблизительно K2O |
|---|---|---|
| Калий хлорид | KCl | от 60 до 62% |
| Сульфат калия | К2SO4 | 50% |
| Сульфат калия-магния (K-mag или Sul-Po-Mag) | K2SO4�2MgSO4 | 20% |
| Тиосульфат калия | К2С2О3 | 17% |
| Нитрат калия | КНО3 | 44% |
Внесение калия и управление им
Предлагаемые методы управления калием различаются для каждой культуры.
Вы можете вносить калийные удобрения осенью или весной для большинства почв в Миннесоте. Песчаные почвы с низкой емкостью катионного обмена обладают низкой способностью удерживать К. Считайте калий частично подвижным на песчаных почвах и вносите его ближе к сроку посева.
|
Таблица 4: Эффективность калийных удобрений при выращивании кукурузы в округе Гудхью
| Коэффициент применения К | Размещение | Урожай зерна |
|---|---|---|
| 0 фунтов. за акр | — | 100 % от максимума |
40 фунтов.
|